

A | B | C | D | E | F | G | H | CH | I | J | K | L | M | N | O | P | Q | R | S | T | U | V | W | X | Y | Z | 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9
Bias in the introduction of variation ("arrival bias") is a theory in the domain of evolutionary biology that asserts biases in the introduction of heritable variation are reflected in the outcome of evolution. It is relevant to topics in molecular evolution, evo-devo, and self-organization.[1][2] In the context of this theory, "introduction" ("origination") is a technical term for events that shift an allele frequency upward from zero (mutation is the genetic process that converts one allele to another, whereas introduction is the population genetic process that adds to the set of alleles in a population with non-zero frequencies). Formal models demonstrate that when an evolutionary process depends on introduction events, mutational and developmental biases in the generation of variation may influence the course of evolution by a first come, first served effect, so that evolution reflects the arrival of the likelier, not just the survival of the fitter.[3][4][5][6][7] Whereas mutational explanations for evolutionary patterns are typically assumed to imply or require neutral evolution, the theory of arrival biases distinctively predicts the possibility of mutation-biased adaptation. [8] Direct evidence for the theory comes from laboratory studies showing that adaptive changes are systematically enriched for mutationally likely types of changes.[9][10][11][12] Retrospective analyses of natural cases of adaptation also provide support for the theory.[11][13] This theory is notable as an example of contemporary structuralist thinking, contrasting with a classical functionalist view in which the course of evolution is determined by natural selection (see [14]).
History
The theory of biases in the introduction process as a cause of orientation or direction in evolution has been explained as the convergence of two threads.[15] The first, from theoretical population genetics, is the explicit recognition by theoreticians (toward the end of the 20th century) that a correct treatment of evolutionary dynamics requires a rate-dependent process of introduction (origination) missing from classical treatments of evolution as a process of shifting frequencies of available alleles.[16][17][18][19] This recognition is evident in the emergence of origin-fixation models that depict evolution as a 2-step process of origination and fixation (by drift or selection), with a rate specified by multiplying a rate of introduction (based on the mutation rate) with a probability of fixation (based on the fitness effect). Origin-fixation models [19] appeared in the midst of the molecular revolution, a half-century after the origins of theoretical population genetics: they were soon widely applied in neutral models for rates and patterns of molecular evolution; their use in models of molecular adaptation was popularized in the 1990s; by 2014 they were described as a major branch of formal theory.[19]
The second thread is a long history of attempts to establish the thesis that mutation and development exert a dispositional influence on evolution by presenting options for subsequent functional evaluation, i.e., acting in a manner that is logically prior to selection. Many evolutionary thinkers have proposed some form of this idea. In the early 20th-century, authors such as Eimer or Cope held that development constrains or channels evolution so strongly that the effect of selection is of secondary importance.[20][21] Early geneticists such as Morgan and Punnett proposed that common parallelisms (e.g., involving melanism or albinism) may reflect mutationally likely changes.[22] Expanding on Vavilov's (1922) [23] exploration of this theme, Spurway (1949) [24] wrote that "the mutation spectrum of a group may be more important than many of its morphological or physiological features."
Similar thinking featured in the emergence of evo-devo, e.g., Alberch (1980) suggests that "in evolution, selection may decide the winner of a given game but development non-randomly defines the players" (p. 665)[25] (see also [26]). Thomson (1985), [27] reviewing multiple volumes addressing the new developmentalist thinking— a book by Raff and Kaufman (1983) [28] and conference volumes edited by Bonner (1982) [29] and Goodwin, et al (1983) [30] — wrote that "The whole thrust of the developmentalist approach to evolution is to explore the possibility that asymmetries in the introduction of variation at the focal level of individual phenotypes, arising from the inherent properties of developing systems, constitutes a powerful source of causation in evolutionary change" (p. 222). Likewise, the paleontologists Elisabeth Vrba and Niles Eldredge summarized this new developmentalist thinking by saying that "bias in the introduction of phenotypic variation may be more important to directional phenotypic evolution than sorting by selection." [31]
However, the notion of a developmental influence on evolution was rejected by Mayr and others such as Maynard Smith ("If we are to understand evolution, we must remember that it is a process which occurs in populations, not in individuals.") [32] and Bruce Wallace ("problems concerned with the orderly development of the individual are unrelated to those of the evolution of organisms through time"), [33] as being inconsistent with accepted concepts of causation. This conflict between evo-devo and neo-Darwinism is the focus of a book-length treatment by philosopher Ron Amundson [34] (see also Scholl and Pigliucci, 2015 [35]). In the theory of evolution as shifting gene frequencies that prevailed at the time, evolutionary causes are "forces" that act as mass pressures (i.e., the aggregate effects of countless individual events) shifting allele frequencies (see Ch. 4 of [36]), thus development did not qualify as an evolutionary cause. A widely cited 1985 commentary on "developmental constraints" [37] advocated the importance of developmental influences, but did not anchor this claim with a theory of causation, a deficiency noted by critics, e.g., Reeve and Sherman (1993) defended the adaptationist program (against the developmentalists and the famous critique of adaptationism by Gould and Lewontin), arguing that the "developmental constraints" argument simply restates the idea that development shapes variation, without explaining how such preferences prevail against the pressure of selection.[38] Mayr (1994) [39] insisted that developmentalist thinking was "hopelessly mixed up" because development is a proximate cause and not an evolutionary one. In this way, developmentalist thinking was received in the 1980s and 1990s as speculation without a rigorous grounding in causal theories, an attitude that persists (e.g., Lynch, 2007 [40]).
.png)







In response to these rebukes, developmentalists concluded that population genetics cannot provide a complete account of evolutionary causation: [41] instead, a dry statistical account of changes in gene frequencies from population genetics must be supplemented with a wet biological account of changes in developmental-genetic organization (called "lineage explanation" in [42]). The beliefs that (1) developmental biology was never integrated into the Modern Synthesis and (2) population genetics must be supplemented with alternative narratives of developmental causation, are now widely repeated in the evo-devo literature and are given explicitly as motivations for reform via an Extended Evolutionary Synthesis. [43]
The proposal to recognize the introduction process formally as an evolutionary cause [3] provides a different resolution to this conflict. Under this proposal, the key to understanding the structuralist thesis of the developmental biologists was a previously missing population-genetic theory for the consequences of biases in introduction. The authors criticized classical reasoning for framing the efficacy of variational tendencies as a question of evolution by mutation pressure, i.e., the transformation of populations by recurrent mutation. They argued that, if generative biases are important, this cannot be because they out-compete selection as forces under the shifting-gene-frequencies theory, but because they act prior to selection, via introduction. Thus the theory of arrival biases proposes that the generative dispositions of a developmental-genetic system (i.e., its tendencies to respond to genetic perturbation in preferential ways) shape evolution by mediating biases in introduction. The theory, which applies to both mutational and developmental biases, addresses how such preferences can be effective in shaping the course of evolution even while strong selection is at work.
Systematic evidence for predicted effects of introduction biases first began to appear from experimental studies of adaptation in bacteria and viruses. [44] [9] Since 2017, this support has widened to include systematic quantitative results from laboratory adaptation, and similar but less extensive results from the retrospective analysis of natural adaptations traced to the molecular level (see below). The empirical case that biases in mutation shape adaptation is considered to be established for practical purposes such as evolutionary forecasting (e.g., [45]). However, the implications of the theory have not been tested critically in regard to morphological and behavioral traits in animals and plants that are the traditional targets of evolutionary theorizing (see Ch. 9 of [15]). Thus, the relevance of the theory to molecular adaptation has been established, but the significance for evo-devo remains unclear. The theory sometimes appears associated with calls for reform from advocates of evo-devo (e.g.,[46][47]), though it has not yet appeared in textbooks or in broad treatments of challenges in evolutionary biology (e.g.,[48][49]).
Simple model
The kind of dual causation proposed by the theory has been explained with the analogy of "Climbing Mount Probable."[50][2] Imagine a robot on a rugged mountain landscape, climbing by a stochastic 2-step process of proposal and acceptance. In the proposal step, the robot reaches out with its limbs to sample various hand-holds, and in the acceptance step, the robot commits and shifts its position. If the acceptance step is biased to favor higher hand-holds, the climber will ascend. But one also may imagine a bias in the proposal step, e.g., the robot may sample more hand-holds on the left than on the right. Then the dual proposal-acceptance process will show both an upward bias due to a bias in acceptance, and a leftward bias due to a bias in proposal. If the landscape is rugged, the ascent will end on a local peak that (due to the proposal bias) will tend to be to the left of the starting point. On a perfectly smooth landscape, the climber will simply spiral to the left until the single global peak is reached. In either case, the trajectory of the climber is subject to a dual bias. These two biases are not pressures competing to determine an allele frequency: they act at different steps, along non-identical dimensions.

The dual effect predicted by the theory was demonstrated originally with a population-genetic model of a 1-step adaptive walk with 2 options,[3] i.e., the climber faces two upward choices, one with a higher selection coefficient and the other with a higher mutation rate. A key feature of model is that neither of the alternatives is present in the initial population: they must be introduced. In simulated adaptation under this model, the population frequently reaches fixation for the mutationally favored allele, even though it is not the most fit option. The form of the model is agnostic with respect to whether the biases are mutational or developmental. Subsequent theoretical work (below) has generalized on the theory of one-step walks, and also considered longer-term adaptive walks on complex fitness landscapes. The general implication for parallel evolution is that biases in introduction may contribute strongly to parallelism. The general implication for the directionality and repeatability of adaptive walks is simply that some paths are more evolutionarily favorable due to being mutationally favorable. The general implication for the long-term predictability of outcomes, e.g., particular phenotypes, is that some phenotypes are more findable than others due to mutational effects, and such effects may strongly shape the distribution of evolved phenotypes.
The application of the theory to problems in evo-devo and self-organization relies formally on the concept of a genotype-phenotype (GP) map. The genetic code, for example, is a GP map that induces asymmetries in mutationally accessible phenotypes. Consider evolution from the Met (amino acid) phenotype encoded by the ATG (codon) genotype. A phenotypic shift from Met to Val requires an ATG to GTG mutation; a shift from Met to Leu can occur by 2 different mutations (ATG to CTG or TTG); a shift from Met to Ile can occur by 3 different mutations (to ATT, ATC, or ATA). If each type of genetic mutation has the same rate, i.e., with no mutation bias per se, the GP map induces 3 different rates of introduction of the alternative phenotypes Val, Leu and Ile. Due to this bias in introduction, evolution from Met to Ile is favored, and this is not due to a mutational bias (in the sense of a bias reflecting the mechanisms of mutagenesis), but rather an asymmetric mapping of phenotypes to mutationally accessible genotypes.
Results of theoretical modeling
One-step adaptive walks
As noted above, in the simplest case of the "Climbing Mount Probable" effect, one may consider a climber facing just two fixed choices: up and to the left, or up and to the right. This case is modeled using simulations by,[3] and is given a more complete treatment by [4] In general, the limiting behavior of evolution as the supply of new mutations becomes arbitrarily small, i.e., as , is called "origin-fixation" dynamics .[19] The origin-fixation approximation for choosing between the left and right options and (respectively) in the Yampolsky-Stoltzfus model is given by the following:



| (1) |

where (or ) and (or ) are the mutation rate and selection coefficient for the left (or right) alternative, and assuming that the probability of fixation . In the Yampolsky-Stoltzfus model, this approximation is good for .






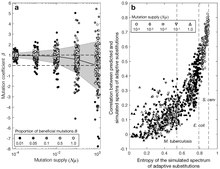


For 1-step walks under origin-fixation conditions, the behavior given by Eqn (1) generalizes from 2 to many alternatives. For instance, Cano, et al. (2022) [7] consider a model gene with many different beneficial mutations, and under low mutation supply, the effects of mutation bias are proportional on the spectrum of adaptive changes.
When is not very small, different beneficial alleles may be present simultaneously, competing and slowing down adaptation, an effect known as clonal interference. Clonal interference reduces the effect of mutation bias in models of evolution in finite genetic spaces: alleles favored by mutation still tend to arrive sooner, but before they reach fixation, later-arising alleles that are more beneficial can out-compete them, enhancing the effect of fitness differences. Under the most extreme condition when all possible beneficial alleles are reliably present in a large population, the most fit allele wins deterministically and there is no room for an effect of mutation bias.[7][51] Stated differently, when all the beneficial alleles are present and selection determines the winner, the chance of success is 1 for the most fit allele, and 0 for all other alleles. Thus, in a gene model with a finite set of beneficial mutations, the influence of mutation bias is expected to be strongest when but to fall off as becomes large.


The influence of mutation under varying degrees of clonal interference can be quantified precisely using the regression method of Cano, et al (2022).[7] Suppose that the expected number of changes of a given class of mutational changes defined by starting and ending states is directly proportional to the product of (1) the frequency of the starting state and (2) the mutation rate raised to the power of , that is,


| (beta1) |

Taking the logarithm of this equation gives